

棕榈酸PA(棕榈酸钠SP)诱导BEECs细胞凋亡、脂质堆积、线粒体功能障碍并增强自噬(IF=6.3)
本文先以梯度浓度的PA(0.2 mmol/L、0.4 mmol/L)作用牛子宫内膜上皮细胞(BEECs)12、24小时,建立细胞损伤模型
过量PA刺激,导致了BEECs线粒体氧化应激和功能障碍,并增强自噬,从而引发了细胞凋亡。MT预处理降低了线粒体氧化应激,并通过AMPK-mTOR-Beclin-1途径进一步增强了受损线粒体的自噬清除,这共同减轻了PA诱导的线粒体功能障碍和BEECs的凋亡。本研究阐明了PA如何对BEECs产生损伤的机制,以及MT如何减轻PA诱导的BEECs损伤,这为预防NEB引起的生殖障碍提供了机制参考。
棕榈酸PA(棕榈酸钠SP)诱导BEECs细胞凋亡、脂质堆积、线粒体功能障碍并增强自噬(IF=6.3)
Journal of Animal Science and Biotechnology [2024](IF=6.3)
1. 前言
负能量平衡(NEB)通常发生在奶牛分娩后。高产奶牛更有可能发生显著的NEB。这种代谢失衡可能导致酮症,并常伴随繁殖性能的下降。然而,NEB的分子机制尚未完全阐明。在过度NEB期间,体内脂肪被大量分解,导致以棕榈酸(PA)为代表的非酯化脂肪酸(NEFAs)在子宫内异常蓄积。这种异常蓄积可能损伤牛子宫内膜上皮细胞(BEECs),而其参与PA诱导BEECs损伤的分子机制仍知之甚少。褪黑素(Melatonin,MT)在维持线粒体活性氧(mitoROS)的稳态中起重要作用。然而,MT是否能改善PA诱导的BEECs损伤及其分子机制尚知之甚少。
2.体外细胞模型构建方法
采用0.2 mmol/L至0.4 mmol/L的PA作用12至24小时可以增加细胞和线粒体的氧化应激水平,如活性氧(ROS)水平增加所示。本文中棕榈酸(PA)源自西安鲲创科技发展有限公司(Kunchuang biotechnology,Xi'an,China)(现货号KC002、旧货号SYSJ-KJ002)。采用可靠的高脂细胞添加剂,能够保证溶剂无毒、无菌、常温无析出、低温无析出、浓度精准,而且无需多次加热助溶,显著减少了操作步骤,提升了实验的稳定性。实际使用时,只需要将高脂细胞添加剂按照比例加入到完全培养基中即可。

3.实验结果
1)PA诱导BEECs细胞凋亡
在复苏和培养后,观察到细胞状况良好。免疫荧光(IF)结果显示,超过99%的细胞表达上皮细胞标志物角蛋白CK18(图1A),表明细胞纯度很高可用于后续实验。我们采用0.2 mmol/L至0.4 mmol/L的PA以建立PA高脂应激模型,PA处理后BEECs脂质含量显著增加,并呈剂量依赖性(Fig.1B,P<0.05)。这些结果证明该模型建立成功。接下来,我们发现细胞活力(图1C,P<0.05)和增殖率(图1D和E,P<0.05)均以剂量依赖性方式显著降低。流式细胞术数据还显示,给予0.2 mmol/L PA处理后存活细胞数显著减少(P<0.05);与对照组相比,早期凋亡和晚期凋亡细胞的比例显著增加(图1F和G,P<0.05)。根据细胞活力、增殖和凋亡数据,我们选择0.2 mmol/L PA处理进行后续实验。然后,我们检测了0.2 mmol/L PA处理后,凋亡相关的mRNA和蛋白的表达。结果显示,PA处理后,促凋亡基因(Caspase3,BAX/BCL2)的mRNA表达显著升高(P<0.05),凋亡相关蛋白(cleaved-Caspase3,BAX)也显著上调(图1H-J,P<0.01)。而BAX/BCL2蛋白的表达水平无差异。总的来说,这些数据表明PA处理诱导了BEECs凋亡。
2)PA增强BEECs的过氧化作用
PA产生的脂毒性诱导的细胞凋亡通常与氧化应激有关。我们发现细胞内和线粒体ROS水平均显著上调(图2A-B和2 E-F,P < 0.05)。此外,在PA刺激下,氧化产物MDA的水平显著增加(图2C,P < 0.01),并且抗氧化剂的活性在PA刺激下显著降低(图2C,P < 0.01)。抗氧化剂活性SOD酶活性显著降低(图2D,P < 0.05)。我们的数据表明,PA诱导的BEECs细胞凋亡增加可能与氧化损伤有关,特别是在线粒体中。
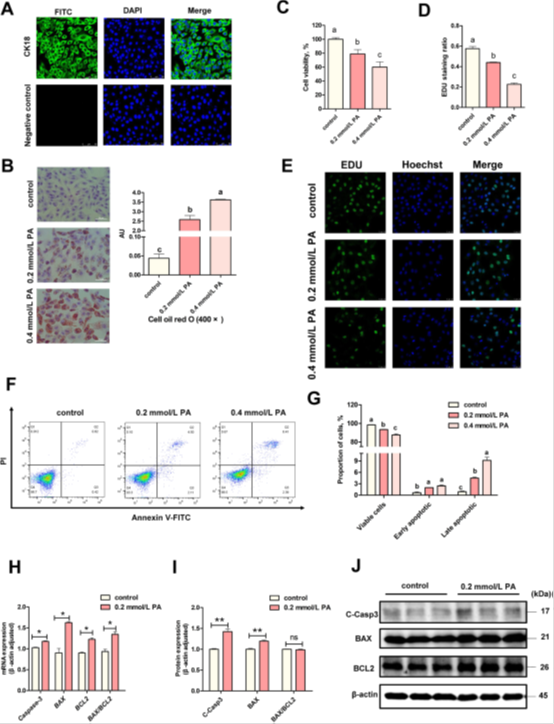
图1 PA诱导BEECs凋亡。(A)BEECs中CK18染色的代表性图像(比例尺=50μm)。(B)油红O染色(×400)的代表性图像和分析(n=3)。(C)在不同浓度的PA刺激下BEEC的细胞活力(n=5)。(D)EDU染色的代表性图像。(E)EDU染色的分析(n=3)。(F,G)用PA刺激BEECs后,通过流式细胞术分析对照组和PA处理组中BEECs的凋亡(n=3)。(H)对照组和PA组(n=3)中BAX、BCL2、Caspase3的qRT-PCR分析。(I,J)对照组和PA组(n=3)中BAX、BCL2、切割的半胱天冬酶3的蛋白质印迹分析。对于B-D、H和I,我们使用单因素方差分析进行显著性差异分析(不同小写字母表示P<0.05);对于H和I,我们使用学生t检验来鉴定显著性差异(*P<0.05;*P<0.01);数据以平均值±SEM显示。
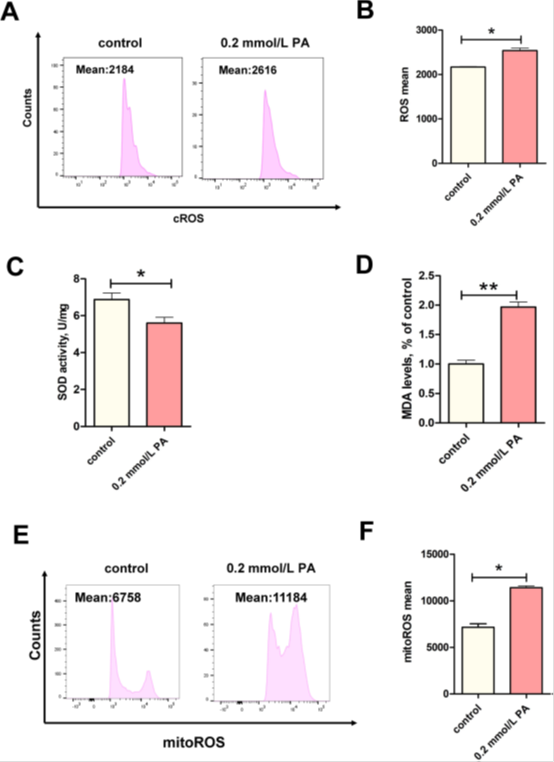
图2 PA诱导BEEC线粒体氧化应激。(A,B)对照组和PA-0.2mmol/L组的cROS水平(n=3)。(C)对照组和PA-0.2 mmol/L组的MDA水平(n=3)。(D)对照组和PA-0.2 mmol/L组的SOD活性(n=5)。(E,F)对照组和PA-0.2 mmol/L组的MitoROS水平(n=3)。对于B-D和F,我们使用Studentt检验确定显著差异(*P<0.05;**P<0.01);数据显示为平均值±SEM。
3)PA诱导BEEC线粒体功能障碍
PA是一种能量代谢底物,线粒体作为细胞内进行脂肪酸氧化的细胞器,在PA应激后,随着线粒体ROS水平升高,BEECs的结构和功能可能受到损害。透射电镜(TEM)显示PA组中的线粒体发生肿胀、嵴溶解和膜破裂(图3A)。统计学分析显示,与对照组相比,受损线粒体的比例显著增加(图3C,P<0.05)。线粒体结构的损伤通常导致线粒体功能障碍,例如MMP的下降,如JC-1染色(图3G)和细胞流式细胞术(图3H和I)所示。提取的线粒体与OXPHOS抗体孵育后,NDUFB8(复合物I,P<0.05)和SDHB(复合物II,P<0.01)蛋白的表达水平显着降低,同时MTCO1(复合物IV)呈下降趋势(图3J-M)),表明PA损害了电子呼吸链的功能。使用Seahorse分析仪进行的OCR分析表明,PA刺激后,线粒体基础细胞呼吸、ATP产生和最大耗氧量均显著下降(图3N-Q,P<0.01),这是线粒体呼吸功能受损的结果。这些观察结果表明,PA诱导的线粒体功能障碍有助于驱动BEECs的损伤。有趣的是,我们发现短杆状线粒体的数量增加,这可能是线粒体分离出来的受损部分。此外,我们通过TEM检测到线粒体数量的减少(图3B,P<0.05),以及线粒体DNA水平的降低(图3D-F),这可能是活跃的自噬引起的。
4)PA激活BEEC中的自噬
为了研究PA是否能激活自噬,我们首先检测了线粒体自噬相关基因(PINK、PARKIN、LC3B、PARL和P62)的mRNA表达,发现它们在0.2 mmol/L PA处理后均显著增加。同时,BEECs中PINK、PARKIN、LC3-II/I和P62的蛋白表达水平也增加,表明更多的线粒体被标记为自噬。我们进一步提取线粒体,以探索PINK、PARKIN、P62和泛素(Ub)的表达,这些蛋白在自噬前期位于或连接到线粒体。P62和Ub蛋白的水平显著增加,而PINK和PARKIN的蛋白水平也略上调(但不显著,P > 0.05),进一步支持我们之前的结果。然后,转染mCherry-GFP-LC3B显示,PA处理后BEECs中的红点显著增加,这初步提示已经形成了更多的自噬小体。上述结果证明自噬前准备工作已充分完成。此外,通过TEM测定,PA处理后BEECs中的溶酶体数量显著增加。LysoTracker染色也表明溶酶体的数量显著增加。PA组除溶酶体数量增加外,PA组具有较强的荧光强度,表明PA显著降低了pH值。LAMP 1和Cat B蛋白表达的增加与我们早期的发现一致。这可能意味着细胞增加了溶酶体的数量和功能,以满足自噬体大量增加的需求。从自噬前后的阶段来看,PA处理后线粒体自噬确实增强。然而,同样值得注意的是,P62蛋白的表达水平显著升高,这可能是自噬增强但未能阻止凋亡的主要原因。
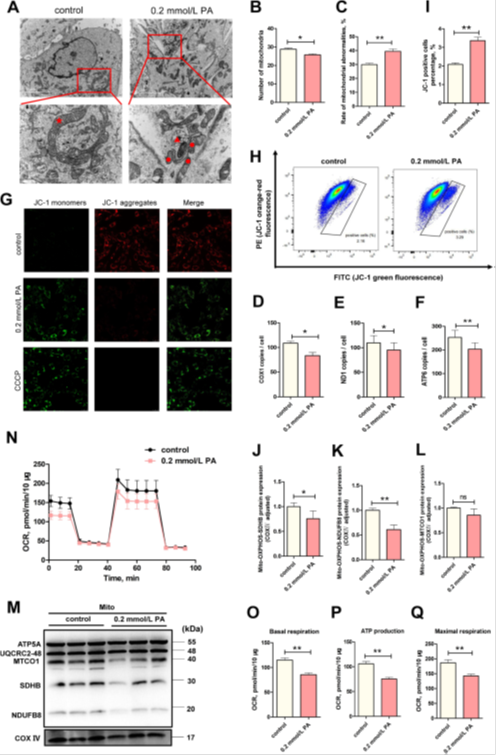
图3 PA导致BEECs中线粒体功能障碍。(A)3组中TEM的代表性图像(比例尺=5和2μm)。正常线粒体用红星标记,异常线粒体用红色圆圈标记,溶酶体用红色三角形标记。(B)TEM图中线粒体平均数的统计分析(n=3)。(C)异常线粒体的TEM统计分析(n=3)。(D-F)线粒体拷贝数分析。采用特异性COX1、ND1和ATP6基因测定法进行测定(n=3)。(G)对照组和PA-0.2 mmol/L组中JC-1染色的代表性图像(n=5)。(H,I)通过流式细胞术测定的JC-1细胞的代表性图像和分析图(n=3)。(J-M)对照组和PA组中OXPHOS的蛋白质印迹分析(n=3)。(N)对照组和PA-0.2 mmol/L组的OCR。(O-Q)通过Seahorse分析仪测定的基础呼吸、ATP生成以及最大呼吸(n=4)。使用学生t检验来鉴定显著差异(*P<0.05;**P<0.01);数据显示为平均值±SEM。
4.总结
本文观察到线粒体功能障碍,包括线粒体结构和呼吸功能的异常,同时线粒体膜电位和线粒体拷贝数减少,并诱导了细胞凋亡。值得注意的是,研究者还观察到自噬相关蛋白(PINK,Parkin,LC3B和Ubiquitin)的上调,然而,P62蛋白也有所增加。100μmol/L MT预处理可抑制PA诱导的线粒体ROS,恢复线粒体呼吸功能。同时,MT预处理可逆转PA诱导的P62蛋白表达上调,激活AMPK-mTOR-Beclin-1通路,促进自噬增加,凋亡减少。
总而言之,过量PA刺激,导致了BEECs线粒体氧化应激和功能障碍,并增强自噬,从而引发了细胞凋亡。MT预处理降低了线粒体氧化应激,并通过AMPK-mTOR-Beclin-1途径进一步增强了受损线粒体的自噬清除,这共同减轻了PA诱导的线粒体功能障碍和BEECs的凋亡。本研究阐明了PA如何对BEECs产生损伤的机制,以及MT如何减轻PA诱导的BEECs损伤,这为预防NEB引起的生殖障碍提供了机制参考。
小编有话说:
本文先以梯度浓度的PA(0.2 mmol/L、0.4 mmol/L)作用牛子宫内膜上皮细胞(BEECs)12、24小时,建立细胞损伤模型。然后,根据细胞活力、增殖和凋亡数据,作者选择0.2 mmol/L PA处理BEECs进行后续实验。上述实验条件和结果,可能会因加药时的细胞密度、代数、状态或耐药性的不同而有所区别。因此,建议具体实验中,先通过浓度、时间梯度实验,寻找出适合于所用细胞的最佳药物作用浓度和最佳作用时间,一般建议在100-500 umol/L范围内、作用24-48小时。
参考文献
Yi Wang, Jianfei Gong, Nuo Heng, et al. " Melatonin alleviates palmitic acid‑induced mitochondrial dysfunction by reducing oxidative stress and enhancing autophagy in bovine endometrial epithelial cells." Journal of Animal Science and Biotechnology.

